ISSN: 1839-9940

 Global reach, higher impact
Global reach, higher impactJ Genomics 2022; 10:61-68. doi:10.7150/jgen.75779 This volume Cite
Research Paper
Draft Genomes of Symbiotic Frankia Strains AgB32 and AgKG'84/4 from Root Nodules of Alnus Glutinosa growing under Contrasted Environmental Conditions
Philippe Normand1 ![]() , Petar Pujic1, Danis Abrouk1, Spandana Vemulapally2, Trina Guerra2, Camila Carlos-Shanley2, Dittmar Hahn2
, Petar Pujic1, Danis Abrouk1, Spandana Vemulapally2, Trina Guerra2, Camila Carlos-Shanley2, Dittmar Hahn2
1. Université Claude-Bernard Lyon 1, Université de Lyon, UMR 5557 CNRS Ecologie Microbienne, Villeurbanne, Cedex 69622, France.
2. Texas State University, Department of Biology, 601 University Drive, San Marcos, TX 78666, USA.
Received 2022-6-3; Accepted 2022-7-8; Published 2022-8-8
Abstract

The genomes of two nitrogen-fixing Frankia strains, AgB32 and AgKG'84/4, were isolated from spore-containing (spore+) and spore-free (spore-) root nodules of Alnus glutinosa, but they did not sporulate upon reinfection. The two strains are described as representatives of two novel candidate species. Phylogenomic and ANI analyses indicate that each strain represents a novel species within cluster 1, with genome sizes of 6.3 and 6.7 Mb smaller than or similar to those of other cultivated Alnus-infective cluster 1 strains. Genes essential for nitrogen-fixation, clusters of orthologous genes, secondary metabolite clusters and transcriptional regulators analyzed by comparative genomic analyses were typical of those from Alnus-infective cluster 1 cultivated strains in both genomes. Compared to other cultivated Alnus-infective strains with large genomes, those of AgB32 and AgKG'84/4 had lost 380 or 409 genes, among which one hup cluster, one shc gene and the gvp cluster, which indicates genome erosion is taking place in these two strains.
Keywords: Frankia, Actinorhizal symbiosis, genome, nitrogen-fixing frankiae, biosynthetic gene clusters
Introduction
Bacteria classified in the genus Frankia constitute a heterologous group of filamentous soil bacteria that can trigger the development of symbiotic root nodules on a range of host plants belonging to 25 genera of perennial, dicotyledonous, angiosperms [1-3]. Isolates have been classified into four distinct clusters, among which three comprise strains that fix atmospheric nitrogen (N2), either in pure culture or in nodules, while cluster 4 frankiae for the most part do not fix N2, except for one strain, and are often unable to fulfil Koch's postulates [4, 5]. Cluster 1 comprises strains infective on Alnus and Casuarina, with currently four described species and two candidate species [6]. The species have type strains deposited in culture collections as Frankia alni ACN14aT [7], F. torreyi CpI1T [8], F. casuarinae CcI3T [7] and F. canadensis ARgP5T [9]. Candidate species represent uncultured Frankia populations in root nodules of host plants, i.e. Candidatus F. nodulisporulans AgTrS, AgUmASt1 and AgUmASH1 [10] and Candidatus F. alpina AiOr, and AvVan [10] that have resisted all attempts at culture.
Several published works on genus Frankia using sub-cluster, OTU, group and genomospecies assignments did provide grounds permitting to affirm that cluster 1 is probably much more diverse than the four species and two candidate species described so far [4, 11-14]. This statement is supported by recent genome analyses of strains Ag45/Mut14 and AgPM24 as representatives of a yet undescribed species [15], and by comparative sequence analyses of amplicons of an actinobacteria-specific insertion in the 23S rRNA genes of frankiae that identified several strains clustering together but that are distinct from type strains of cluster 1 [16]. Strains AgB32 and AgKG'84/4 are two such strains, isolated from root nodules of Alnus glutinosa growing under contrasted environmental conditions at two locations in Germany about 350 km apart. Strain AgB32 was isolated from spore[+] root nodules of Alnus glutinosa of a forest ecotype that was interspersed with oak (Quercus robur) in an established riverside forest on a wet, but well aerated sandy loam in Bad Bentheim, Germany (52.320319, 7.159997) [17]. Strain AgKG'84/4 was isolated from spore[-] root nodules of A. glutinosa of the pioneer ecotype growing in a pure stand at a lake shore marsh in water-logged soil rich in organic material in Krems II-Goels, Germany (53.989103, 10.360772) [17]. Both strains had previously been identified as members of cluster 1, representing a subcluster designated as subgroup I [18] or cluster 1d [16]. In order to assess the viability of the previous amplicon-based analysis and to potentially amend and refine the species diversity of cluster 1 frankiae, we used whole genome sequence analyses trying to affirm the potential of strains AgB32 and AgKG'84/4 for the description of new species.
Materials and Methods
Sample preparation
Frankia strains AgB32 and AgKG'84/4 were grown from stocks preserved in 20% vol/vol glycerol at -80 °C since 2003 in Defined Propionate Medium (DPM) containing propionate and NH4Cl as C and N source, respectively (19), at 30 °C for two weeks. Cells were harvested by centrifugation (15,000 × g, 5 min) and homogenized through sonication (10 s at 20% output in a S-450 sonifier, Branson Ultrasonics, Danbury, CT) [20]. DNA was extracted from cell pellets after an additional centrifugation step using the SurePrepTM Soil DNA Isolation Kit (Fisher Scientific, Houston, TX) [21], and concentrations measured with a Qubit® 2.0 Fluorometer (Life Technologies, Carlsbad, USA). Library preparation and sequencing using the Illumina tagmentation protocol and the NextSeq Illumina platform (2 × 150 bp) using standard protocols were done at the Microbial Genomics Sequencing Center, Pittsburgh, PA, USA.
Genome assembly
Default settings of fastp were used to filter and trim sequence reads [22], with reads with average %GC<54 removed using bbduk (https://jgi.doe.gov/data-and-tools/bbtools/bb-tools-user-guide/). SPAdes 3.13.0 was used to assemble genomes [23] and QUAST to check their quality [24]. Genome completeness was estimated using the lineage workflow (lineage_set) CheckM v1.0.18 with default values [25].
Comparative genomic analysis
Assembled genomes of strains AgB32 and AgKG'84/4 as well as Frankia genomes of type strains of all described species and other selected genomes were selected for Average Nucleotide Identity (ANI) comparisons [26] using the pyani platform with the b (Blast) setting ([27]; https://pyani.readthedocs.io). Genomes were further analyzed on the Mage platform [28] to compute clusters of orthologous genes (COGs) [29], to identify secondary metabolite clusters through antiSMASH [30] and to identify genes specific to or lost in the new genomes. A MASH distance matrix [31] was used to construct a phylogenetic tree using a rapid neighbour joining algorithm [32] on the Mage platform.
Results
Characteristics of the two Frankia genomes
The genomes of the two strains AgB32 and AgKG'84/4 were considered complete given their CheckM scores of 99.59% and 98.05%, respectively. The N50 were 55 309 and 112 139, respectively and the total length were 6 667 069 and 6 426 475. They were considered pure with contamination indices of 1.09 and 2.37, respectively. Genomes of AgB32 and AgKG'84/4 harbored 214 and 1,305 contigs with the largest contig being 223 506 nt and 54 816 nt, respectively. Their GC contents of 72.23 and 71.88% for AgB32 and AgKG'84/4, respectively (Table 1).
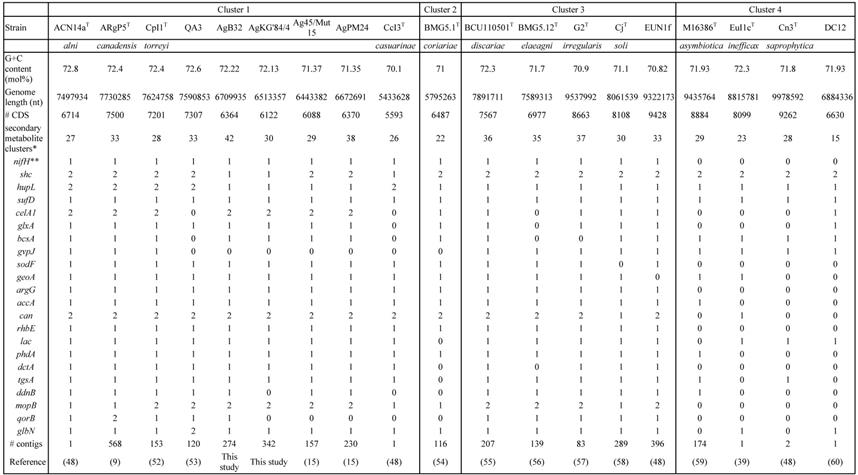
Basic genome characteristics (G+C%, genome length, number of CDS, number of secondary metabolite clusters, presence of selected genes, # of contigs and references) of Frankia strains AgB32 and AgKG'84/4 compared to those of type strains of Frankia species in clusters 1 to 4
Phylogenetic analysis of Frankia spp
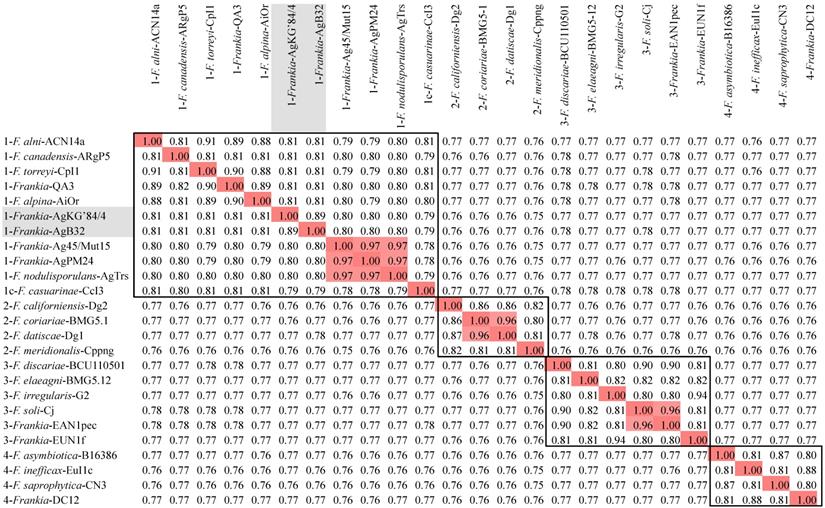
A phylogenetic tree generated from the MASH matrix with Frankia genomes of type strains revealed that the closest strains to AgB32 and AgKG'84/4 were members of cluster 1 (Figure 1). Average nucleotide identity (ANI) between strains AgB32 and AgKG'84/4 was 89%, indicating that they belong to two separate genospecies (Figure 2). ANI values at or below 80% were obtained for both strains in comparison with Frankia genomes of type strains of all described species (Figure 2). The ANI values with other cluster 1 genomes ranged from 79% (CcI3) to 81% (ACN14a), while 76-77% values were obtained with cluster 2 genomes, and 77-78% with cluster 3 and 4 genomes (Figure 2).
Phylogenetic tree based on comparative sequence analyses of complete genomes of Frankia species and candidate species, using Streptomyces coelicolor (AL645882) as outgroup. Frankia clusters 1 to 4 are indicated on the right. Scale units are substitutions per site. The two genomes described in the present study have red arrows.

Heatmap matrix of Average Nucleotide Identity (ANI) comparisons (in percent) for the Frankia genomes of type strains of described species using the pyani platform with the b (Blast) setting [27]; https://pyani.readthedocs.io). The two genomes described in the present study are highlighted in grey. Those ANI values above the 95% threshold are highlighted in red. ANI values of clusters are boxed.
Analysis of functional genes in Frankia spp. isolates
All genes identified as playing a role in the symbiosis were found to be present in the genomes of AgB32 and AgKG'84/4, i.e. nif, hup, suf, shc, cel, glx, bcsA (Table 1). Furthermore, all genes that are more abundant in symbiotic lineages (clusters 1, 2 and 3) than in non-symbiotic lineages (cluster 4) (sodF, geoA, argF, accA, rhbE, dctA, phdA, tgsA, ddnB) were also recovered in AgB32 and AgKG'84/4 (Table 1). Conversely, gvp that codes for gas vesicle proteins, one of the two shc genes and one of the two hup clusters that are found in cluster 1 strains were not found in the two genomes while the symbiotic cluster was maintained [33].
The COG computation showed values for AgB32 and AgKG'84/4 characteristic of other Alnus-infective cluster 1 strains with a low number of categories “N” (Cell motility), and “P” (Inorganic ion transport and metabolism) (Table 2). These results are similar for the antiSMASH computation that showed AgB32 and AgKG'84/4 to have values characteristic of other Alnus-infective cluster 1 strains with a high number of T1PKS and NRPS (Table 3). T1PKS and NRPS typically code for antibiotics and a high number of such clusters is evocative of a good capacity for keeping other soil microbes at bay. The numbers of transcriptional regulators were on the whole comparable to other strains with a low number of ArsR, and LuxR regulators (Table 4).
COG characteristics of Frankia strains AgB32 and AgKG'84/4 compared to those of type strains of Frankia species in clusters 1 to 4
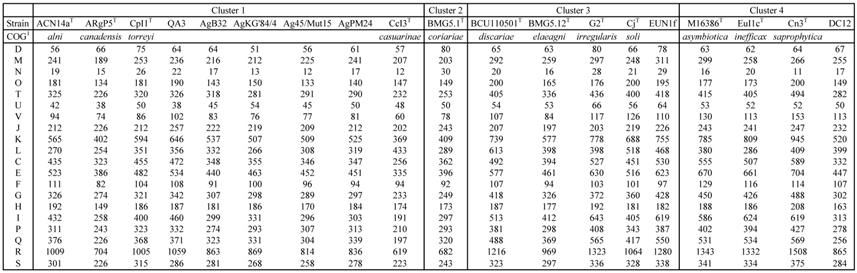
1class: D: Cell cycle control, cell division, chromosome partitioning; M: Cell wall/membrane/envelope biogenesis; N: Cell motility; O: Posttranslational modification, protein turnover, chaperones; T: Signal transduction mechanisms; U: Intracellular trafficking, secretion, and vesicular transport; V: Defense mechanisms; J: Translation, ribosomal structure and biogenesis; K: Transcription; L: Replication, recombination and repair; C: Energy production and conversion; E: Amino acid transport and metabolism; F: Nucleotide transport and metabolism; G: Carbohydrate transport and metabolism; H: Coenzyme transport and metabolism; I: Lipid transport and metabolism; P: Inorganic ion transport and metabolism; Q: Secondary metabolites biosynthesis, transport and catabolism; R: General function prediction only; S: Function unknown.
Number of secondary metabolites clusters (antiSMASH) of Frankia strains AgB32 and AgKG'84/4 compared to those of cultivated type strains of Frankia species in clusters 1 to 4
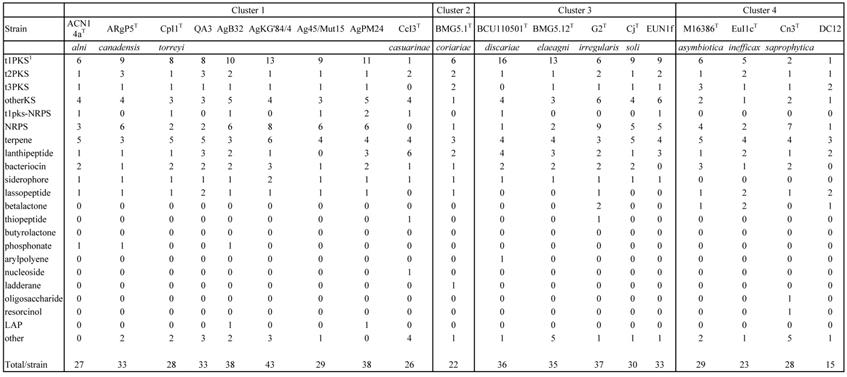
1tnPKS is type “n” Polyketide Synthase;
NRPS is Non Ribosomal Peptide Synthase, LAP is Linear Azole/azoline-containing Peptide.
Number of transcriptional regulators of Frankia strains AgB32 and AgKG'84/4 compared to those of type strains of Frankia species in clusters 1 to 4
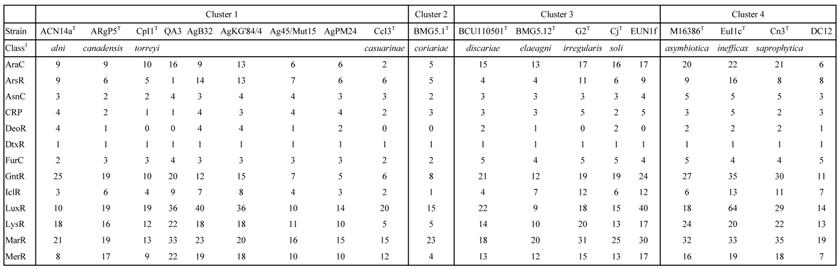
1class: AraC: arabinose regulator; ArsR: arsenic resistance; AsnC: asparagine synthase regulator; CRP: cyclic AMP receptor protein (catabolite repression); DeoR: deoxyribonucleoside synthesis operon regulator; DtxR: diphtheria toxin repressor; FurC: ferric uptake regulator; GntR: gluconate regulator; IclR: isocitrate lyase regulator; LuxR: quorum-sensing luminescence regulator; LysR: lysine regulator; MarR: Multiple antibiotic resistance regulator; MerR: mercury resistance regulator; TetR: Tetracycline repressor; WhiB: regulation of morphological differentiation.
A search for genes present in F. alni ACN14a, Frankia sp. QA3, F. torreyi CpI1 and F. canadensis ARgP5 but absent in AgB32 and AgKG'84/4 yielded 380 or 409 hits, respectively among which an alkane sulfonate, a acetyl/propionyl CoA carboxylase locus, an uptake hydrogenase locus, a dicarboxylate transporter, a Hup locus, the GVP locus, several transporters (Table S1). Conversely, there were 565 genes present in both AgB32 and AgKG'84/4 but absent in F. alni ACN14a, Frankia sp. QA3, F. torreyi CpI1 and F. canadensis ARgP5, of which about half (277) were of unknown function.
Discussion
The genus Frankia has been scantily described for many years because of difficulties to isolate and grow frankiae in pure culture, a major prerequisite for the description of strains [34, 35]. Some populations to this day have even resisted isolation attempts so far [36]. Differentiation of isolates has also been hampered by the availability of few distinguishing features between populations [14]. Starting in 2007, new developments in whole genome sequencing techniques have overcome these difficulties and resulted in the determination of genome sequences of three Frankia isolates [37], and ultimately even of uncultured Frankia populations in root nodules [38]. Comparative analyses of whole genome sequences between Frankia populations have resulted in the description of twelve species and five candidate species for uncultured populations so far [6]. These numbers were based on the availability of 37 genomes [39], a number that is increasing regularly [15, 40]. Comparative sequences analyses of whole genomes and metrics such as ANI [26] or dDDH [41] are now used as foundation for the description of microbial genera, species and subspecies.
Members of the genus Frankia have been assigned into four clusters, numbered 1 to 4, within the genus [4]. These assignments have proven quite solid over the years, with cluster 1 in particular found to remain coherent with all Alnus-infective symbiotic strains. Cluster 1c with Casuarina-infective strains remains at the root of this cluster with several distinguishing features such as the lack of vesicles in nodules, a host-derived hemoglobin protection against oxygen and a distinct host range [6]. Alnus-infective symbiotic strains have been described initially on the basis of DNA/DNA homology as quite close to one another [14] but the full extent of diversity has slowly emerged with studies targeting new cultured strains and uncultured frankiae from specific environments [38, 42-47].
Genomes of Alnus-infective symbiotic strains have initially been found to be quite large at 7.5 Mb with several ancient duplicated genes such as the shc gene coding for the synthesis of hopanoid lipids [48], the hup genes coding for hydrogen uptake for the recycling of hydrogen derived from nitrogenase [33], the cel coding for cellulases [49], the can coding for the carbonic anhydrase necessary for feeding short chain fatty acids (SCFA) into the tricarboxylic acid (TCA) cycle or the kor genes coding for 2-oxoglutarate ferredoxin oxidoreductase that connects the TCA cycle with nitrogenase (with the nitrogen-fixation process) [50, 51]. Some of these duplications have been found to be lost in lineages with smaller genome size as is the case for Ag45/Mut15 and AgPM24 [15]. It appears the genomes of strains AgB32 and AgKG'84/4 are also undergoing a parallel process of genome erosion. This process is similar with some of the genes lost in common such as hup but also other genes such as shc only lost in AgB32 and AgKG'84/4.
AgB32 and AgKG'84/4 are two distinct lineages with an ANI of 89%, well below the threshold of 95 set by Goris [26] to delineate species but markedly above the 80% average between other Alnus-infective cluster 1 species. This would indicate the two strains should constitute two distinct species yet sharing many features due to a recent common ancestry.
Supplementary Material
Supplementary table.
Acknowledgements
The authors are indebted to the Graduate College (Doctoral Research Support Fellowship to S. Vemulapally), and the Department of Biology at Texas State University for financial support.
Competing Interests
The authors have declared that no competing interest exists.
References
1. Schwintzer CR, Tjepkema JD. The biology of Frankia and actinorhizal plants, Academic Press, San Diego, 1990
2. Huss-Danell K. Actinorhizal symbioses and their N-2 fixation. New Phytol. 1997;136:375-405
3. Benson DR, Silvester WB. Biology of Frankia strains, actinomycete symbionts of actinorhizal plants. Microbiol Rev. 1993;57:293-319
4. Normand P, Orso S, Cournoyer B, Jeannin P, Chapelon C, Dawson J. et al. Molecular phylogeny of the genus Frankia and related genera and emendation of the family Frankiaceae. Int J Syst Bacteriol. 1996;46:1-9
5. Gtari M, Nouioui I, Sarkar I, Ghodhbane-Gtari F, Tisa LS, Sen A. et al. An update on the taxonomy of the genus Frankia Brunchorst, 1886, 174AL. Anton Leeuw. 2019;112:5-21
6. Normand P, Fernandez MP. Frankia Brunchorst 1886, 174AL. In: Whitman WB, Rainey FA, Kämpfer P, Trujillo ME, DeVos P, Hedlund B, Dedysh S. (Eds.) Bergey's Manual of Systematics of Archaea and Bacteria. 2019 10.1002/9781118960608.gbm00042.pub2
7. Nouioui I, Ghodhbane-Gtari F, Montero-Calasanz MD, Goker M, Meier-Kolthoff JP, Schumann P. et al. Proposal of a type strain for Frankia alni (Woronin 1866) Von Tubeuf 1895, emended description of Frankia alni, and recognition of Frankia casuarinae sp nov and Frankia elaeagni sp nov. Int J Syst Evol Microbiol. 2016;66:5201-5210
8. Nouioui I, Ghodhbane-Gtari F, Jando M, Tisa LS, Klenk HP, Gtari M. Frankia torreyi sp. nov, the first actinobacterium of the genus Frankia Brunchorst 1886, 174AL isolated in axenic culture. Anton Leeuw. 2019;112:57-65
9. Normand P, Nouioui I, Pujic P, Fournier P, Dubost A, Schwob G. et al. Frankia canadensis sp nov, isolated from root nodules of Alnus incana subspecies rugosa. Int J Syst Evol Microbiol. 2018;68:3001-3011
10. Herrera-Belaroussi A, Normand P, Pawlowski K, Fernandez MP, Wibberg D, Kalinowski J. et al. Candidatus Frankia nodulisporulans sp. nov, an Alnus glutinosa-infective Frankia species unable to grow in pure culture and able to sporulate in-planta. Syst Appl Microbiol. 2020;43(6):126134
11. Hahn D. Polyphasic taxonomy of the genus Frankia. In: Pawlowski K, Newton WE (eds), Nitrogen-fixing actinorhizal symbioses. Springer Verlag, Berlin, Germany. 2007 pp. 25-48
12. Hahn D, Mirza B, Benagli C, Vogel G, Tonolla M. Typing of nitrogen-fixing Frankia strains by matrix-assisted laser desorption ionization-time-of-flight (MALDI-TOF) mass spectrometry. Syst Appl Microbiol. 2011;34:63-68
13. Pozzi AC, Bautista-Guerrero HH, Abby SS, Herrera-Belaroussi A, Abrouk D, Normand P. et al. Robust Frankia phylogeny, species delineation and intraspeciesdiversity based on Multi-Locus Sequence Analysis (MLSA) and Single-Locus Strain Typing (SLST) adapted to a large sample size. Syst Appl Microbiol. 2018;41:311-323
14. Fernandez MP, Meugnier H, Grimont PAD, Bardin R. Deoxyribonucleic acid relatedness among members of the genus Frankia. Int J Syst Bacteriol. 1989;39:424-429
15. Normand P, Pujic P, Abrouk D, Vemulapally S, Guerra T, Carlos-Shanley C. et al. Draft genomes of nitrogen-fixing Frankia strains Ag45/Mut15 and AgPM24 isolated from root nodules of Alnus glutinosa. Journal of Genomics. 2022 10.7150/jgen.74788
16. Ben Tekaya S, Ganesan AS, Guerra T, Dawson JO, Forstner MRJ, Hahn D. SybrGreen- and TaqMan-based quantitative PCR approaches allow assessment of the abundance and relative distribution of Frankia clusters in soils. Appl Environ Microb. 2017;83(5):e02833-16
17. Hahn D, Dorsch M, Stackebrandt E, Akkermans ADL. Synthetic oligonucleotide probes for identification of Frankia strains. Plant Soil. 1989;118:211-219
18. Samant S, Huo T, Dawson JO, Hahn D. Abundance and relative distribution of Frankia host infection groups under actinorhizal Alnus glutinosa and non-actinorhizal Betula nigra trees. Microb Ecol. 2016;71:473-481
19. Meesters TM, van Genesen ST, Akkermans ADL. Growth, acetylene reduction activity and localization of nitrogenase in relation to vesicle formation in Frankia strains Cc1.17 and Cp1.2. Arch Microbiol. 1985;143:137-142
20. Mirza BS, Welsh A, Hahn D. Saprophytic growth of inoculated Frankia sp. in soil microcosms. FEMS Microbiol Ecol. 2007;62:280-289
21. Samant S, Sha Q, Iyer A, Dhabekar P, Hahn D. Quantification of Frankia in soils using SYBR Green based qPCR. Syst Appl Microbiol. 2012;35:191-197
22. Chen S, Zhou Y, Chen Y, Gu J. fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics. 2018;34:i884-i890
23. Prjibelski A, Antipov D, Meleshko D, Lapidus A, Korobeynikov A. Using SPAdes De Novo Assembler. Curr Protoc Bioinformatics. 2020;70:e102
24. Gurevich A, Saveliev V, Vyahhi N, Tesler G. QUAST: quality assessment tool for genome assemblies. Bioinformatics. 2013;29:1072-1075
25. Parks DH, Imelfort M, Skennerton CT, Hugenholtz P, Tyson GW. CheckM: assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015;25:1043-1055
26. Goris J, Konstantinidis KT, Klappenbach JA, Coenye T, Vandamme P, Tiedje JM. DNA-DNA hybridization values and their relationship to whole-genome sequence similarities. Int J Syst Evol Microbiol. 2007;57:81-91
27. Pritchard L, Glover RH, Humphris S, Elphinstone JG, Toth IK. Genomics and taxonomy in diagnostics for food security: soft-rotting enterobacterial plant pathogens. Anal Methods-Uk. 2016;8:12-24
28. Vallenet D, Calteau A, Cruveiller S, Gachet M, Lajus A, Josso A. et al. MicroScope in 2017: an expanding and evolving integrated resource for community expertise of microbial genomes. Nucleic Acids Res. 2017;45:D517-D528
29. Tatusov RL, Natale DA, Garkavtsev IV, Tatusova TA, Shankavaram UT, Rao BS. et al. The COG database: new developments in phylogenetic classification of proteins from complete genomes. Nucleic Acids Res. 2001;29:22-28
30. Medema MH, Blin K, Cimermancic P, de Jager V, Zakrzewski P, Fischbach MA. et al. antiSMASH: rapid identification, annotation and analysis of secondary metabolite biosynthesis gene clusters in bacterial and fungal genome sequences. Nucleic Acids Res. 2011;39:W339-346
31. Ondov BD, Treangen TJ, Melsted P, Mallonee AB, Bergman NH, Koren S. et al. Mash: fast genome and metagenome distance estimation using MinHash. Genome Biol. 2016;17:132
32. Simonsen M, Mailund T, Pedersen CNS, Rapid Neighbour-Joining, in: K.A. Crandall, J. Lagergren (Eds.) WABI 2008: Algorithms in Bioinformatics, Springer Verlag, Heidelberg, 2008, pp. 113-122
33. Richau KH, Kudahettige RL, Pujic P, Kudahettige NP, Sellstedt A. Structural and gene expression analyses of uptake hydrogenases and other proteins involved in nitrogenase protection in Frankia. J Biosciences. 2013;38:703-712
34. Lechevalier MP. Taxonomy of the genus Frankia (Actinomycetales). Int J Syst Bacteriol. 1994;44:1-8
35. Lechevalier MP, Lechevalier HA, Systematics, isolation, culture of Frankia. In: Schwintzer CR, Tjepkema JD (eds), The biology of Frankia and actinorhizal plants. Academic Press, New York. 1990 pp. 35-60
36. Cotin-Galvan L, Pozzi AC, Schwob G, Fournier P, Fernandez MP, Herrera-Belaroussi A. In-planta sporulation capacity enhances infectivity and rhizospheric competitiveness of Frankia strains. Microbes Environ. 2016;31:11-18
37. Normand P, Queiroux C, Tisa LS, Benson DR, Rouy Z, Cruveiller S. et al. Exploring the genomes of Frankia. Physiol Plantarum. 2007;130:331-343
38. Bethencourt L, Vautrin F, Taib N, Dubost A, Castro-Garcia L, Imbaud O. et al. Draft genome sequences for three unisolated Alnus-infective Frankia Sp+ strains, AgTrS, AiOr and AvVan, the first sequenced Frankia strains able to sporulate in-planta. J Genomics. 2019;7:50-55
39. Tisa LS, Oshone R, Sarkar I, Ktari A, Sen A, Gtari M. Genomic approaches toward understanding the actinorhizal symbiosis: an update on the status of the Frankia genomes. Symbiosis. 2016;70:5-16
40. Carlos-Shanley C, Guerra T, Hahn D. Draft genomes of non-nitrogen-fixing Frankia strains. J Genomics. 2021;9:68-75
41. Auch AF, von Jan M, Klenk HP, Goker M. Digital DNA-DNA hybridization for microbial species delineation by means of genome-to-genome sequence comparison. Stand Genomic Sci. 2010;2:117-134
42. Welsh AK, Dawson JO, Gottfried GJ, Hahn D. Diversity of Frankia in root nodules of geographically isolated Arizona alders in central Arizona (USA). Appl Environ Microbiol. 2009;75:6913-6918
43. Welsh A, Mirza BS, Rieder JP, Paschke MW, Hahn D. Diversity of frankiae in root nodules of Morella pensylvanica grown in soils from five continents. Syst Appl Microbiol. 2009;32:201-210
44. Mirza BS, Welsh A, Rasul G, Rieder JP, Paschke MW, Hahn D. Variation in Frankia populations of the Elaeagnus host infection group in nodules of six host plant species after inoculation with soil. Microb Ecol. 2009;58:384-393
45. Pozzi AC, Bautista-Guerrero HH, Nouioui I, Cotin-Galvan L, Pepin R, Fournier P. et al. In-planta sporulation phenotype: a major life history trait to understand the evolution of Alnus-infective Frankia strains. Environ Microbiol. 2015;17:3125-3138
46. Pozzi ACM, Herrera-Belaroussi A, Schwob G, Bautista-Guerrero HH, Bethencourt L, Fournier P. et al. Proposal of 'Candidatus Frankia alpina', the uncultured symbiont of Alnus alnobetula and A. incana that forms spore-containing nitrogen-fixing root nodules. Int J Syst Evol Microbiol. 2020;70:5453-5459
47. Ben Tekaya S, Guerra T, Rodriguez D, Dawson JO, Hahn D. Frankia diversity in host-plant root nodules is independent of abundance or relative diversity of Frankia in corresponding rhizosphere soils. Appl. Environ. Microbiol. 2018;84:e02248-17
48. Normand P, Lapierre P, Tisa LS, Gogarten JP, Alloisio N, Bagnarol E. et al. Genome characteristics of facultatively symbiotic Frankia sp strains reflect host range and host plant biogeography. Genome Res. 2007;17:7-15
49. Pujic P, Alloisio N, Fournier P, Roche D, Sghaier H, Miotello G. et al. Omics of the early molecular dialogue between Frankia alni and Alnus glutinosa and the cellulase synton. Environ Microbiol. 2019;21:3328-3345
50. Dodsworth JA, Cady NC, Leigh JA. 2-oxoglutarate and the PII homologues NifI(1) and NifI(2) regulate nitrogenase activity in cell extracts of Methanococcus maripaludis. Mol Microbiol. 2005;56:1527-1538
51. Huergo LF, Dixon R. The emergence of 2-Oxoglutarate as a master regulator metabolite. Microbiol Mol Biol Rev. 2015;79:419-435
52. Oshone R, Hurst SGt, Abebe-Akele F, Simpson S, Morris K, Thomas WK. et al. Permanent draft genome sequences for two variants of Frankia sp. strain CpI1, the first Frankia strain isolated from root nodules of Comptonia peregrina. Genome Announc. 2016;4(1):e01588-15
53. Sen A, Beauchemin N, Bruce D, Chain P, Chen A, Walston Davenport K. et al. Draft genome sequence of Frankia sp. strain QA3, a nitrogen-fixing actinobacterium isolated from the root nodule of Alnus nitida. Genome Announc. 2013;1:e0010313
54. Gtari M, Ghodhbane-Gtari F, Nouioui I, Ktari A, Hezbri K, Mimouni W. et al. Cultivating the uncultured: growing the recalcitrant cluster-2 Frankia strains. Sci Rep-Uk. 2015;5:13112 doi: 13110.11038/srep13112
55. Wall LG, Beauchemin N, Cantor MN, Chaia E, Chen A, Detter JC. et al. Draft genome sequence of Frankia sp. strain BCU110501, a nitrogen-fixing actinobacterium isolated from nodules of Discaria trinevis. Genome Announc. 2013;1(4):e00503-13
56. Nouioui I, Beauchemin N, Cantor MN, Chen A, Detter JC, Furnholm T. et al. Draft genome sequence of Frankia sp. strain BMG5.12, a nitrogen-fixing actinobacterium isolated from Tunisian soils. Genome Announc. 2013;1(4):e00468-13
57. Nouioui I, Gtari M, Goker M, Ghodhbane-Gtari F, Tisa LS, Fernandez MP. et al. Draft genome sequence of Frankia strain G2, a nitrogen-fixing actinobacterium isolated from Casuarina equisetifolia and able to nodulate actinorhizal plants of the order Rhamnales. Genome Announc. 2016;4(3):e00437-16
58. Gtari M, Ghodhbane-Gtari F, Nouioui I. Frankia soli sp. nov, an actinobacterium isolated from soil beneath Ceanothus jepsonii. Int J Syst Evol Microbiol. 2020;70:1203-1209
59. Nouioui I, Gueddou A, Ghodhbane-Gtari F, Rhode M, Gtari M, Klenk HP. Frankia asymbiotica sp nov, a non-infective actinobacterium isolated from Morella californica root nodule. Int J Syst Evol Microbiol. 2017;67:4897-4901
60. Tisa LS, Beauchemin N, Cantor MN, Furnholm T, Ghodhbane-Gtari F, Goodwin L. et al. Draft genome sequence of Frankia sp. strain DC12, an atypical, noninfective, ineffective isolate from Datisca cannabina. Genome Announc. 2015;3(4):e00889-15
Author contact
![]() Corresponding author: Philippe Normand, Université Claude-Bernard Lyon 1, Université de Lyon, UMR 5557 CNRS Ecologie Microbienne, Villeurbanne, Cedex 69622, France. E-mail: philippe.normandfr.
Corresponding author: Philippe Normand, Université Claude-Bernard Lyon 1, Université de Lyon, UMR 5557 CNRS Ecologie Microbienne, Villeurbanne, Cedex 69622, France. E-mail: philippe.normandfr.